|
А.С. Степановских
Экология. Учебник для вузов
М.: ЮНИТИ-ДАНА, 2001. - 703 с.
10. Внутривидовые и межвидовые взаимоотношения в популяциях, гомеостаз и экологические стратегии
10.1. Внутривидовые взаимоотношения
Многообразное
население популяции постоянно взаимодействует между собой. Удовлетворение
потребностей в питании, распределении кормовых угодий, выбор места для постройки
гнезда, спаривание, выращивание потомства, охрана занимаемой территории,
расселение и т. д. осуществляются при постоянном взаимодействии особей,
входящих в каждую популяцию, которая и обеспечивает ее существование.
Эти
связи складывались по мере образования и развития вида как целостной системы.
Поэтому все особи, входящие в популяцию, обладают и общностью происхождения, и
многочисленными специфическими приспособлениями к совместной жизни (рис. 10.1).
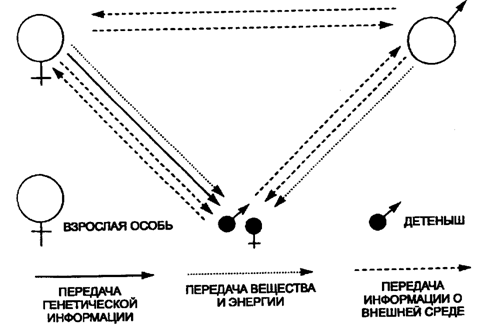
Рис. 10.1. Связи особей в популяции (на
примере млекопитающих)
Эти
приспособления, названные С.А. Северцовым (1951) конгруэнциями,
охватывают морфофизиологические и этологические (поведенческие) черты. Сюда
можно отнести: особенности строения, обеспечивающие встречи разнополых особей,
размножение, выращивание молодняка, приспособления, обеспечивающие расселение
или объединения в стаи (к миграциям или на зиму), разнообразные «сигналы» —
запахи, цвет, голос, поведение и др. — все, что привлекает или отвлекает
особей, предупреждает их о занятой территории (рис. 10.2).

Рис.
10.2. Информационная нагрузка разных поз собак
Данные
приспособления могут носить характер индивидуальных и групповых контактов. Они
по-разному осуществляются на разных стадиях развития организмов и
могут меняться в течение жизни особи, в разные сезоны года, а также в связи с
изменениями условий жизни.
Взаимоотношения
между членами популяции зависят прежде всего от того, одиночный или групповой
образ жизни свойствен виду. Формы же существования особей в популяции чрезвычайно
различны.
Одиночный
образ жизни. Особи популяции
обособлены и независимы друг от друга. Характерен для многих видов, главным
образом на определенных стадиях жизненного цикла. Полностью одиночное
существование организмов в природе не встречается. Причиной этого является
невозможность осуществления их основной жизненной функции — размножения. Однако
для некоторых видов характерны очень слабые контакты между совместно живущими
особями. Так, у видов с внутренним оплодотворением встречи самцов и самок могут
быть очень кратковременными, для осуществления копуляции. В остальное время
животные живут отдельно, независимо друг от друга, например хищные
жуки-жужелицы, божьи коровки и многие другие насекомые. Подобный образ жизни
ведут отдельные водные обитатели (одиночные актинии) с нарушенным способом
оплодотворения, при котором нет необходимости в непосредственной встрече
партнеров. Нередко виды с одиночным образом жизни образуют временные скопления
особей в период, предшествующий размножению, в местах зимовок и т. д.
Бабочки-крапивницы поздней осенью в большом количестве собираются в чердачных
помещениях, божьи коровки и жужелицы — возле пней и комлей деревьев в сухой
подстилке, щуки и сомы — в зимовальных ямах на дне водоема. Вместе с тем
подобные скопления не сопровождаются установлением тесных связей между
животными. Каждое из них относительно независимо от остальных.
Усложнение
отношений внутри популяций происходит по двум направлениям: усилению связи
между половыми партнерами и возникновению контактов между родительским и
дочерними поколениями. В популяциях на этой основе формируются семьи,
разнообразные по составу и длительности существования. Родительские пары могут
создаваться на короткий или длительный срок, а у некоторых видов — на всю жизнь
взрослых особей. Среди птиц тетерева, глухари не образуют устойчивых семейных
пар. У многих воробьиных самка и самец держатся вместе в течение всего периода
гнездования. Сохраняются на долгие годы семейные пары лебедей, журавлей,
голубей. Выбор партнеров у животных сопровождается особым брачным поведением,
нередко большой сложности, — «танцы», «ухаживания» и др.
У
многих насекомых, птиц и млекопитающих ухаживание нередко предотвращает
агрессивные и оборонительные реакции особей противоположного пола, приводит к
синхронизации полового созревания, стимулирует готовность к спариванию, что
имеет большое значение для размножения, к которому самец и самка должны быть
готовы в одно и то же время.
В
период выбора половых партнеров в популяциях животных усиливаются конкурентные
отношения. У многих животных возникают драки самцов, ритуальные демонстрации и
другие типы специализированного поведения, которые направлены на устранение
конкурентов (рис. 10.3).
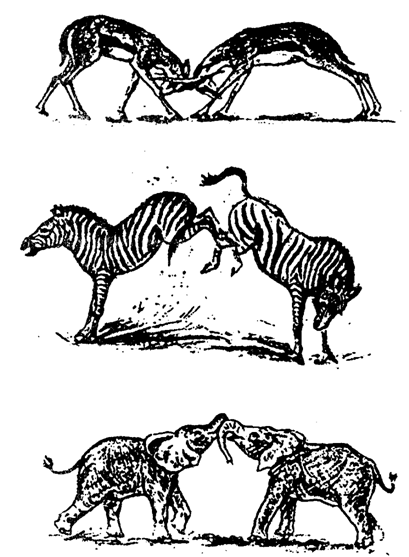
Рис.
10.3. Турнирные бои в «брачный период» у
млекопитающих (по А. О. Рувинскому и др., 1993)
Несмотря
на частую ожесточенность, эти столкновения редко приводят к большим травмам
соперников, большей частью ограничиваются изгнанием одного из них с территории,
где находится самка. Токующие туруханы стремительно бросаются друг на друга, а
затем внезапно замирают со взъерошенным «воротником», после этого вновь
повторяют броски. В таких сражениях у рыб преобладает ритуал угрозы или
противники кусают друг друга в пасть — наименее уязвимую часть тела, но не
наносят более опасных укусов в бок.
Следовательно,
период, предшествующий размножению в популяциях животных, характеризуется
активным поиском и резким усилением контактов между особями.
Семейный
образ жизни. Усиливает связи между
родителями и их потомством. Простейшим видом такой связи является, например,
забота одного из родителей об отложенных яйцах — охрана кладки, инкубации,
дополнительное аэрирование и т. д. У птиц забота о птенцах продолжается до
поднятия их на крыло, а у ряда крупных млекопитающих, таких, как медведи,
тигры, детеныши воспитываются в семейных группах до наступления их половой
зрелости, в течение нескольких лет. В зависимости от того, кто из родителей
берет на себя уход за потомством, различают семьи отцовского, материнского
и семейного типов. В охране и выкармливании потомства в семьях с
устойчивым образованием пар принимают участие обычно как самец, так и самка.
Дальнейшее
усложнение поведенческих связей в популяциях приводит к формированию более
крупных объединений животных — колоний, стай, стад.

Рис. 10.4. Колониальные гнездовья у птиц
(из Н. М. Черновой, А. М.
Быловой, 1981):
А
— колонии белых гусей в тундре;
В
— колонии грачей
Колонии — групповые
поселения оседлых животных, которые могут существовать длительное время или создаваться
на период размножения, как у птиц (гуси, грачи, гагары, чайки и др.), рис.
10.4.
Колонии
животных отличаются разнообразием — от простых территориальных скоплений
одиночных форм до объединений, где отдельные члены, как органы в целостном
организме, выполняют разные функции видовой жизни. Так, сифонофора Salacia
— единая, на первый взгляд, особь — образована множеством отдельных
специализированных особей и представляет колонию.
Развитие
колонии начинается с одного индивидуума, который размножается почкованием.
Отдельные отпочковывающиеся особи могут либо вести самостоятельную жизнь, либо
стать специализированными частями материнской колонии.
Поселения
животных, где некоторые функции их жизни выполняются сообща, что увеличивает
вероятность выживания отдельных особей,является более сложной формой колонии.
Такими общими функциями колонии чаще всего становятся защита от врагов и
предупредительная сигнализация. Чайки, ласточки, гуси и другие птицы с шумом
набрасываются на хищника, угрожающего кладкам или птенцам. Поднятая заметившей
опасность птицей тревога мобилизует остальных. Птицам сообща удается изгонять
крупных хищников, с которыми они не справились бы поодиночке — сов, ястребов,
песцов и др. Часто сохраняются индивидуальные гнездовые участки в колониальных
поселениях птиц. Так, у серебристых чаек колонии рыхлые, между гнездами
сохраняется расстояние 3—5 м.
Городские
же ласточки лепят свои гнезда часто вплотную одно к другому. Территориальные
инстинкты здесь не проявляются совсем. Общественные ткачики строят на деревьях
большое общее гнездо из травы с многочисленными отверстиями, ведущими в
индивидуальные гнездовые полости.
Колонии
млекопитающих (сурки, вискачи, пеструшки, пищухи) чаще возникают на основе
разрастания семейных групп, с сохранением связей между отпочковывающимися
семьями.
Сложные
колонии у общественных насекомых, таких, как муравьи, термиты, пчелы, возникают
на основе сильно разрастающейся семьи. В колониях-семьях насекомые выполняют
сообща большинство основных функций: размножения, защиты, обеспечения кормом
себя и потомства, строительств» и т. д. Здесь существует обязательное
разделение труда и специализация отдельных особей, возрастных групп на
выполнение определенных операций. Члены колонии действуют на основе постоянного
обмена информацией друг с другом.
По
мере усложнения колониального объединения поведение, а нередко физиология и
строение отдельной особи все больше и больше подчиняются интересам всей
колонии.
Стаи — временные
объединения животных, которые проявляют биологически полезную организованность
действий. Стаи облегчают выполнение каких-либо функций в жизни вида: добыча
пищи, защита от врагов, миграции. Стайность наиболее широко распространена
среди рыб и птиц, у млекопитающих — многих собачьих. В стаях сильно развиты
подражательные реакции и ориентация на соседей.
Действия
стаи по способам координации действий делятся на две категории: ) эквипотенциальные
— без выраженного доминирования отдельных членов; 2) стаи с лидерами,
где животные ориентируются на поведение одной или нескольких, как правило,
наиболее опытных особей. Объединения первого типа известны у рыб, мелких птиц,
перелетной саранчи. У крупных птиц и млекопитающих встречается обычно второй
тип стай.
Стаи
рыб изменчивы по величине, форме, плотности, нередко переформировываются по
нескольку раз в сутки. Рыбы группируются в стаи в светлое время суток, при
зрительном контакте с другими особями, и рассредоточиваются на ночь. Защитная
роль стайных объединений рыб велика. При опасности стая рыб быстро маневрирует,
обтекая хищника, который, например, бросившись в ее середину, оказывается в
пустыне. Для поведения рыб в стае характерен имитационный рефлекс — подражание
действиям соседей.
Стаи
у птиц формируются при сезонных перелетах или у оседлых и кочующих форм, при
зимних кормежках. При перелетах стаи образуют те виды, которым свойственно
колониальное гнездование или коллективное кормление. Одиночно гнездящиеся и
кормящиеся виды стай в полете не образуют.
Существует
постоянная сигнализация, звуковая и зрительная связь между особями в стаях
оседлых птиц, которая используется для обнаружения благоприятных мест ночевок и
отдыха, источников корма, восходящих потоков воздуха и т. д. Так, подобно
пионерам Дикого Запада в вестернах, виргинские перепелки образуют на ночь
кольцо «спина к спине» (рис. 10.5). Такая привычка не только помогает им
уберечься от хищников, но и согревает холодными осенними и зимними ночами.
Вспугнутые перепелки врассыпную взлетают в разные стороны.

Рис. 10.5 Кольца
перепелок (по П. Фарбу, 1971)
Для
групповой охоты зимой возникают волчьи стаи. Им удается стаей справиться с
крупными копытными, на которых охота в одиночку часто безрезультатна. При
групповой охоте волки обычно практикуют преследование с выходом на перехват
жертвы, нагон жертвы на засаду или захват ее в кольцо, требующее
согласованности и координации действий всех особей (рис. 10.6).

Рис. 10.6. Охота
волков на северных оленей
(по Н. М.
Черновой и др., 1995)
В
стаях млекопитающих значительна роль вожаков и специфичны отношения между
отдельными особями, что сближает данные групповые образования со стадами.
Стада — более
длительные и постоянные объединения животных по сравнению со стаями. Здесь
осуществляются все основные функции жизни вида: добывание корма, защита от хищников,
миграции, размножение, воспитание молодняка и т. д. В основе группового
поведения животных в стадах лежат взаимоотношения доминирования-подчинения,
основанные на индивидуальных различиях между особями. Одним из вариантов
организации стад являются группы с временными или относительно постоянными
лидерами. Это особи, на которых концентрируется внимание других, а они, в свою
очередь, своим поведением определяют направление перемещения, места кормления,
реакцию на хищника и т. д. Деятельность лидера не направлена непосредственно на
подчинение других особей. Лидером становится более опытный член стада. Стадо
действует как единое целое, подражая лидеру. Так, стада северных оленей, как
правило, ведут старые важенки. Они лучше других ориентируются при миграциях и
нападении хищников, потому что периодически это им приходится делать в
одиночку.
Выделяются
объединения в крупных стадах, которые представляют семейные или возрастные
группы с внутренними, более дружелюбными контактами, чем с членами других
аналогичных групп. Независимо от общего лидерства во внутривидовых группировках
могут складываться отношения администрирования-подчинения.
Наибольшей
сложностью отличается поведенческая организация стад с вожаками и иерархическим
соподчинением особей. В отличие от лидеров вожаки характеризуются поведением,
направленным непосредственно на активное руководство стадом: специальными
сигналами, угрозами и прямыми нападениями. Здесь нередко возникают разделения
«прав» и «обязанностей» и более сложные формы общественного поведения, выгодные
для группы в целом. В стаде ранг каждой особи определяется многими причинами:
возрастом, физической силой, опытом и наследственными качествами животного. Как
правило, над слабыми доминируют сильные и опытные, с устойчивым типом нервной системы.
Это проявляется в праве на самку, преимуществе при поедании пищи, передвижении
в группе и др.
Доминирование-подчинение
весьма различно у разных видов. Главные из них — «линейная» иерархия типа
«треугольника», деспотия. При линейной иерархии в ряду рангов А-В-С и т.
д. особи, принадлежащие к каждому, подчинены предыдущим, но главенствуют над
последующими. В таком ряду последние животные — самые бесправные в группе. Так,
вожаки в стаях ездовых собак активно подчиняют себе всю стаю, угрожая и задавая
трепку непослушным. Животные низшего ранга ведут себя покорно перед всеми
остальными, подходят к пище в последнюю очередь. Они изгоняются с лучших мест
отдыха, не подпускаются к самкам и т. д.
У
некоторых животных иерархическое соподчинение осуществляется по типу «треугольника»:
А нападает на В, В — на С, а С подчиняет себе А. Данное соотношение может
сохраняться в группе довольно долго. Следующим вариантом иерархии является деспотия,
или доминирование одного животного над всеми остальными членами группы.
Ранг
животного в группе определяется столкновениями между особями в виде прямой
борьбы или ритуальных угроз. После установления ранга всех членов группы прямые
столкновения между ними прекращаются и порядок поддерживается сигнальным или
ритуальным поведением. Иерархически организованному стаду свойствен
закономерный порядок перемещения, определенная организация при защите,
расположении на местах отдыха и т. д. Например, когда начинают маячить хищники,
самки слонов образуют круг вокруг детенышей, чтобы охранить их от нападающих
(рис. 10.7).

Рис. 10.7. Самки слонов охраняют
детенышей от хищников
В
стаде павианов в наибольшей безопасности, в центре, находятся самки,
готовящиеся к размножению или с детенышами, по краям — вожаки, молодые самцы и
неразмножающиеся самки. Впереди и позади стада шествуют крупные самцы, готовые
отразить нападение. Известны случаи, когда при преследовании хищниками стада
самец-доминант возвращался за отставшим детенышем, хотя ему грозила большая
опасность.
Биологический
смысл иерархической системы доминирования-подчинения заключается в создании
согласованного поведения группы, выгодного для всех ее членов. После
«расстановки сил» животные не тратят лишней энергии на индивидуальные
конфликты, а в целом группа получает преимущества, подчиняясь наиболее сильным
и опытным индивидуумам. Это имеет большое значение для выращивания молодняка,
обеспечения защиты от хищников, предупреждения от опасности, миграциях и т. д.
Например, в сложных ситуациях (голодовках и др.) гибнут большей частью слабые,
подчиненные особи, но под защитой группы они имеют больше возможности выжить,
чем в одиночку. Иерархия четко выражена не только в стадах у млекопитающих, но
и в колониях у птиц, у ряда беспозвоночных: насекомых (сверчков,
жуков-чернотелок и др.), некоторых ракообразных и пр.
Эффект
группы. Многие виды животных, как уже
отмечалось, нормально развиваются только тогда, когда объединяются в довольно
большие группы. Например, бакланы (Phalacrocarax bougainvillei) могут существовать в колонии, которая насчитывает не менее 10000 особей
и где на 1 м3 приходится не менее 3 гнезд. Жизнь в группе через
нервную и гормональную системы отражается на протекании многих физиологических
процессов в организме животного. Наблюдается тесное общение особей посредством
запахов, звуков, специфики поведения. Благодаря сложной системе сигнализации у
особей и их взаимному обмену информацией возрастает эффективность
функционирования группы, направленная на удовлетворение важных жизненных
потребностей всех ее членов. Оптимизация физиологических процессов, ведущая
к повышению жизнеспособности при совместном существовании, и получила название
«эффект группы». Эффект группы проявляется как психофизиологическая реакция
отдельной особи на присутствие других особей своего вида. Например, у овец вне
стада учащаются пульс и дыхание. При виде приближающегося стада эти процессы
нормализуются. Эффект группы проявляется в ускорении темпов роста животных,
повышении плодовитости, более быстром образовании условных рефлексов, повышении
средней продолжительности жизни индивидуума и др. Животные в группе обычно
способны поддерживать оптимальную температуру, например, при окучивании, в
гнездах, в ульях. Вне группы у многих я животных не реализуется плодовитость.
Не
менее важным показателем эффекта группы служит фазовая изменчивость. Она
была впервые обнаружена Б. П. Уваровым в 1921 г. у саранчовых, а позднее и у
жесткокрылых, чешуекрылых и других насекомых. У саранчовых четко различают две
формы: одиночная и стадная (рис. 10.8).
Главной
причиной здесь является разная плотность особей в популяции. Большая
скученность их и обусловливает образование стадной формы. Особи обеих форм
отличаются окраской, поведением, скоростью развития, строения. Каждой форме
свойственна определенная активность. Особи в стадной форме очень подвижны и
тяготеют к миграциям. Это является средством регуляции численности саранчи в ее
резервациях. Скученный образ жизни в нарастающем темпе приводит к сокращению
яйцевых трубок. Отсюда, чем выше степень стадности, тем ниже плодовитость, т.е.
плодовитость у стадных саранчовых обратно пропорциональна плотности популяции.
Изолированные самки азиатской саранчи откладывают 1000—1200 яиц, а находящиеся
в сильной скученности — только 300. Однако у последних масса отрождающихся
личинок, их выживаемость и общая жизнеспособность значительно выше. Эффект
группы и высшее ее проявление у саранчовых — фазовая изменчивость —
функционально связаны с плотностью популяции и служат механизмом регуляции их
численности. Взаимная стимуляция особей вызывает формирование стадной формы,
которая характеризуется снижением плодовитости, сокращением смертности в ранних
возрастах, увеличением скорости развития и повышением активности.
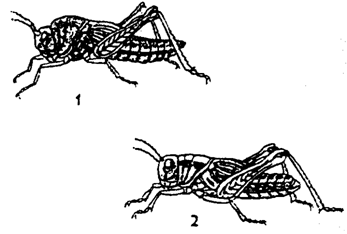
Рис.
10.8. Формы перелетной саранчи:
1 –
одиночная; 2 - стандная
Вспышки
численности грызунов прекращаются большей частью из-за таких эффектов
скученности, как повышенная агрессивность вследствие усиленной секреции
адреналина, вялости, связанная с понижением сахара в крови, чем из-за нехватки
пищи или инфекций. Помет, секреты, продукты обмена веществ могут портить
имеющиеся достаточные запасы питания. Высокая плотность иногда приводит к
каннибализму даже у видов в норме чисто растительноядных, например, таких, как
мучной хрущак. Таким образом, положительный эффект группы проявляется до
некоторого оптимального уровня плотности популяции. Когда животных становится
слишком много, это грозит для всех недостатком ресурсов Среды, вступают в
действие другие механизмы, которые приводят к снижению численности особей в
группе путем ее деления, рассредоточения или падения рождаемости. Эти механизмы
в дальнейшем рассмотрим более подробно.
Агрессия,
внутривидовой паразитизм и конкуренция. Агрессия — это форма связей,
характеризующаяся истреблением особей своего вида. Среди внутривидовых связей
довольно часто встречаются различные формы агрессии, когда истребление одних
особей вида другими способствует поддержанию численности популяции, ее
плотности на занимаемой территории и обеспечивает высокую жизненность
сохранившихся особей. В отдельных случаях агрессия представлена каннибализмом,
или пожиранием особей своего вида. Так, у американской
саламандры-скрытножаберника отложенную икру охраняет самец. Защищая ее от
других, он сам питается ею для поддержания сил. Другая форма каннибализма у
лососевых. Взрослые рыбы для икрометания заходят в реки и обычно поднимаются до
самых верхних истоков. На мелководьях, в условиях хорошо аэрируемой воды, они
выметывают икру и там же обессиленные погибают. Икрометание происходит поздно
осенью и на мелководье, трупы рыб не уносятся потоком воды, а вмерзают в лед.
Весной с таянием льда оттаивают и трупы рыб, начинается их разложение. К этому
времени из икры обычно уже появляется молодь. Прожорливые мальки набрасываются
на размягченные трупы рыб и поедают их. Это позволяет большинству мальков
выживать и быстро расти в местах, где иной доступной им пищи нет. В целом
каннибализм широко распространен у рыб. У трески, налима, балхашского окуня
взрослые рыбы поедают свою молодь. Аналогичное наблюдается и у насекомых.
Каннибализм отмечен у муравьев, у хищных личинок некоторых комаров. Во всех
приведенных случаях агрессия оказывается полезной виду, являя своеобразный
пример «взаимополезных связей».
Таким
же полезным можно оценить явление внутривидового паразитирования,
которое встречается у некоторых животных. Оно может быть в виде эктопаразитизма
(наружного паразитизма) и эндопаразитизма (внутреннего паразитизма).
Внутривидовой
эктопаразитизм особенно четко выражен
у глубоководной рыбы-удильщика. В качестве приспособления для обеспечения
оплодотворения икры самка постоянно носит на себе самца. Карликовый самец
(размер самца 1,5—2 см, самки — 9—10 см) еще в стадии молоди прикрепляется к
самке: у одних видов — к особому отростку на жаберной крышке, у других — к
брюшку или ко лбу (рис. 10.9).
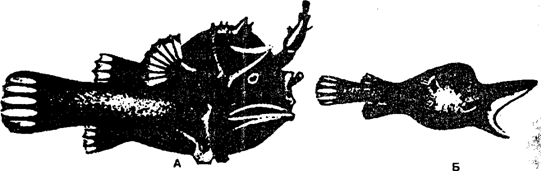
Рис. 10.9. Глубоководная рыба-удильщик:
а
— самка; б — карликовый самец (по Н. М. Черновой и др., 1995)
Кожа
самца в местах прикрепления срастается с кожей самки, как бы погружаясь в нее,
происходит взаимопроникновение и некоторое срастание кровеносных сосудов обоих
организмов. Становясь неотъемлемой частью самки, самец и передвигается за счет
ее. При этом у самца кишечник недоразвит, зубы редуцированы, тогда как жабры,
почки, сердце и половая система остаются достаточно развитыми.
Внутривидовой
эндопаразитизм наблюдается, например,
у бо-неллии — кольчатого червя, живущего в Средиземном море. Самец является
внутренним паразитом самки, поселяясь у нее в канальцах нефридиев, по которым
проходят выводимые наружу яйца. Еще на стадии личинок самец попадает в рот, а
затем в пищевод и позднее через ткани достигает нефридиев, где и остается жить.
Тот и другой случаи внутривидового паразитизма возникли в условиях бедности
кормов, при очень низкой плотности популяции, как приспособление к восстановлению
потомства, где встреча самцов и самок происходит довольно редко. Переход к
паразитированию у этих видов обычно осуществляется во время личиночных стадий,
когда только что выведшиеся мальки и личинки еще держатся одной общей стайкой.
При сравнительно высокой плотности молодых особей в таких условиях
встречаемость самца и самки происходит часто. Этот и целый ряд других видовых
особенностей позволяют виду обеспечить свое воспроизведение и длительное
существование в борьбе с другими видами.
Внутривидовая
конкуренция (за пищу, полового
партнера, жизненное пространство, место для размножения и др.) увеличивается
как с ростом плотности популяции, так и степени специализации вида. Чаще всего
конкуренция начинается за пищу, когда в результате размножения при еще
достаточном запасе пищи плотность популяции повышается. Недостаточное питание
может приводить нередко к снижению плодовитости, пока уменьшение популяции не
позволит виду снова размножиться.
У
растений внутривидовая конкуренция нередко проявляется в виде «пассивной
борьбы». Пассивная борьба приводит к появлению особых адаптивных черт в
строении, обеспечивающих им выгодное размещение своих органов. Это особенно
четко было продемонстрировано В. Н. Сукачевым (1945) в опытах при загущенных
посевах. Было показано, что не только надземные части растений разместились на
разной высоте, но и их корни распределились на разной глубине (рис. 10.10).
Дифференциация
молодых особей наблюдается при загущенном посеве настурции. Среди одновременно
появившихся ее всходов вскоре можно выделить три^ более или менее отличные друг
от Друга группы молодых растений. У одних наблюдается быстрый рост стебля,
выносящий семядольные и первые зеленые листья вверх. У другой группы быстрый
рост вверх осуществляется не за счет роста стебля, а при помощи быстрого роста
черешков семядольных и первых зеленых листьев. У третьей группы не отмечается
быстрого роста ни стеблевой части, ни черешков, но оказалась высокой скорость
развертывания листьев. К тому времени, когда растения первых двух групп имели,
помимо семядольных, лишь два зеленых листа, у этих было развернуто четыре, а в
одном случае даже шесть зеленых листьев с очень мелкими листовыми пластинками.
Во время цветения взрослые растения внешне выглядят одинаково и практически
ничем не напоминают того разделения особей на три группы, которое наблюдается у
проростков.

Рис. 10.10. Надземная и подземная
ярусность в травянистых
фитоценозах (по В. Г.
Хржановскому и др., 1994)
Территориальность. Территориальное поведение встречается у широкого
круга животных, в том числе у некоторых рыб, рептилий, птиц, млекопитающих и
общественных насекомых. Это явление основано на врожденном стремлении особи к
свободе передвижения на некоторой минимальной площади (рис. 10.11).
Первая
ступень развития территориальности — индивидуальное пространство,
окружающее каждую особь. Оно хорошо заметно, например, у ласточек, усевшихся на
телефонный провод, или у скворцов в летящей стае. Особь защищает его от
вторжения и открывает для другой особи только после церемоний ухаживания перед
спариванием. Вторая ступень — обороняемое место для жизни, отдыха или сна в
середине необороняемой зоны активности (у многих хищников охотничьего
участка). Животные, стоящие на второй ступени, распределяются практически
равномерно.
Рис. 10.11.
Территориальное поведение животных
(по
Н. М. Черновой и др., 1995)
Самое
рациональное использование пространства отмечается на третьей ступени
территориальности, где образуются настоящие территории — участки, из
которых другие особи изгоняются. Владелец участка психологически господствует
на нем, и для изгнания в большинстве случаев достаточно лишь демонстраций,
угроз, преследования, самое большое — притворных атак, которые прекращаются на
границах участка, помеченных зрительно, акустически или запахом (ольфакторно)
рис. 10.12.

Рис. 10.12.
Бурый медведь, маркирующий дерево
(по Н.
М. Черновой и др., 1995)
У
птиц, гнездящихся колониями, особь охраняет только свое гнездо, а вся колония и
ее окрестности как целое обороняются всей популяцией.
|
