|
Т.А. Акимова, A.П. Кузьмин, В.В. Хаскин
Экология. Природа - Человек - Техника
Учебник для вузов. – М.: ЮНИТИ-ДАНА, 2001. - 343 с.
Глава 3. Биота биосферы
3.3. Экосистемы
Популяция
или часть популяции какого-либо вида растений или животных может входить в
состав разных сообществ, где она сочетается с различными наборами
представителей других видов. Многовидовые сообщества организмов в природе -
биоценозы (см. стр. 31) - образованы не случайным сочетанием видов, а таким их
подбором, такими количественными соотношениями и такой
пространственно-временной организацией, которые обеспечивают непрерывный
круговорот веществ и четко распределенные потоки энергии. Этим достигается
устойчивость экосистем.
Состав
м функциональная структура экосистемы. Каждая экосистема имеет собственное материально-энергетическое
обеспечение и определенную функциональную структуру, основанную на пищевых (трофических)
взаимоотношениях (рис. 3.2). Эта структура составлена несколькими группами
организмов, каждая из которых выполняет определенную работу в круговороте
веществ. Организмы, относящиеся к одному такому звену, образуют трофический
уровень, а последовательные связи между трофическими уровнями образуют цепи
питания, или трофические цепи. В экосистему входят организмы,
различаемые по способу питания - автотрофы и гетеротрофы.
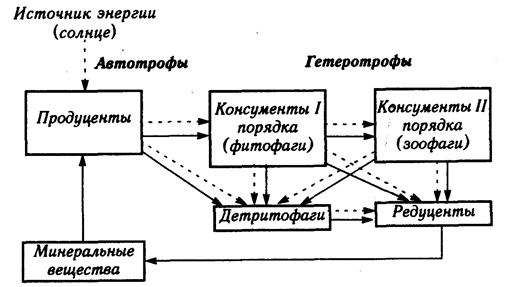
Рис.
3.2. Упрощенная схема переноса веществ и энергии в экосистеме
  - перенос веществ, - перенос энергии - перенос веществ, - перенос энергии
Автотрофы (самопитающие)
- организмы, образующие органическое вещество своего тела из неорганических
веществ - в основном из углекислого газа и воды - посредством процессов
фотосинтеза и хемосинтеза. Фотосинтез осуществляют фотоавтотрофы - все
хлорофиллоносные (зеленые) растения и микроорганизмы. Хемосинтез наблюдается у
некоторых почвенных и водных хемоавтотрофных бактерий, которые используют в
качестве источника энергии не солнечный свет, а ферментативное окисление ряда
веществ - водорода, серы, сероводорода, аммиака, железа.
Фотоавтотрофы
(растения) составляют основную массу биоты и полностью отвечают за образование
всего нового органического вещества в экосистеме, т.е. являются первичными
производителями продукции - продуцентами экосистем. Синтезированная
автотрофами новая биомасса органического вещества - это первичная продукция,
а скорость ее образования - биологическая продуктивность экосистемы.
Автотрофы образуют первый трофический уровень любой полночленной экосистемы.
Гетеротрофы (питающиеся
другими) - организмы, потребляющие готовое органическое вещество других
организмов и продуктов их жизнедеятельности. Это все животные, грибы и большая
часть бактерий.
В
отличие от автотрофов-продуцентов гетеротрофы выступают как потребители и
деструкторы (разрушители) органических веществ. В зависимости от источников
питания и участия в деструкции они подразделяются на консументов и редуцентов.
Консументы - потребители органического вещества организмов. К ним
относятся:
- консументы
I порядка - растительноядные животные (фитофаги), питающиеся живыми
растениями (тля, кузнечик, гусь, овца, олень, слон);
- консументы
II порядка - плотоядные животные (зоофаги), поедающие других животных, -
различные хищники (хищные насекомые, насекомоядные и хищные птицы, хищные
рептилии и звери), нападающие не только на фитофагов, но и других хищников.
Существует немало животных со смешанным питанием, потребляющих и растительную и
животную пищу - плотоядно-растительноядные и всеядные. Консументы I и II
порядка занимают соответственно второй, третий, а иногда и следующий
трофические уровни в экосистеме.
Особенную
группу консументов составляют паразиты и симбиотрофы. И те и
другие живут (по меньшей мере на протяжении части жизненного цикла) за счет
веществ организма-хозяина. Это уже не только животные (черви, насекомые,
клещи), но и различные микроорганизмы, а также некоторые грибы и растения. В
отличие от паразитов, часто вызывающих заболевания, а иногда и гибель хозяина,
симбиотрофы выполняют жизненно важные для хозяина трофические функции. Это
мицелиальные грибы-микоризы, участвующие в корневом питании многих растений;
клубеньковые бактерии бобовых, связывающие молекулярный азот; микробиальное
население сложных желудков жвачных животных, повышающее перевариваемость и
усвоение поедаемой растительной пищи.
Еще
одну группу консументов образуют детритофаги, или сапрофаги -
животные, питающиеся мертвым органическим веществом - остатками и продуктами
жизнедеятельности растений и животных. Это различные черви, членистоногие
(клещи, многоножки, личинки насекомых, жуки-копрофаги) и другие животные - все
они выполняют функцию очищения экосистем. Детритофаги участвуют в образовании
почвы, торфа, донных отложений водоемов.
Редуценты - бактерии и низшие грибы - завершают деструктивную
работу консументов и сапрофагов, доводя разложение органики до ее полной
минерализации и возвращая в среду экосистемы молекулярный азот, минеральные
элементы и последние порции двуокиси углерода.
Все
названные группы организмов в любой экосистеме тесно взаимодействуют между
собой, согласуя потоки вещества и энергии. Их совместное функционирование не
только поддерживает структуру и целостность биоценоза, но и оказывает существенное
влияние на абиотические компоненты биотопа, формируя и поддерживая
экологическую среду экосистемы.
Никакая
часть экосистемы не может существовать без другой. Если по какой-либо причине
происходит нарушение структуры экосистемы, исчезает группа организмов, вид, то
по закону цепных реакций может сильно измениться или даже разрушиться все
сообщество. Но часто бывает и так, что через какое-то время после исчезновения
одного вида на его месте оказываются другие организмы, другой вид, но
выполняющий сходную функцию в экосистеме. Эта закономерность называется правилом,
замещения, или дублирования:
у
каждого вида в экосистеме есть «дублер». Такую роль обычно выполняют виды менее
специализированные и в то же время экологически более гибкие, адаптивные. Так,
копытных в степи замещают грызуны; на мелководных озерах и болотах аистов и цапель
замещают кулики и т.п. При этом решающую роль играет не систематическое
положение, а близость экологических функций между замещаемыми и замещающими
группами организмов.
Принципиальное
различие между потоками вещества и энергии в экосистеме заключается в том, что
биогенные элементы, составляющие органическое вещество, могут многократно
участвовать в круговороте веществ, тогда как поток энергии однонаправлен и
необратим. Каждая порция энергии используется однократно. В соответствии со
вторым законом термодинамики на каждом этапе трансформации энергии значительная
ее часть неизбежно рассеивается в виде теплоты.
Пищевые цепи и трофические уровни. Прослеживая пищевые взаимоотношения между членами
биоценоза («кто кого и сколько поедает»), можно построить пищевые цепи питания
различных организмов. Примером длинной пищевой цепи может служить
последовательность обитателей арктического моря: «микроводоросли (фитопланктон) ® мелкие растительноядные ракообразные (зоопланктон) ® плотоядные планктонофаги (черви, ракообразные, моллюски, иглокожие) ® рыбы (возможны 2-3 звена последовательности хищных рыб) ® тюлени ® белый медведь».
Цепи наземных экосистем обычно короче. Пищевая цепь, как правило, искусственно
выделяется из реально существующей пищевой сети - сплетения многих цепей
питания.
Благодаря
определенной последовательности пищевых отношений различаются отдельные
трофические уровни переноса веществ и энергии в экосистеме, связанные с питанием
определенной группы организмов. Совокупности трофических уровней различных
экосистем моделируются с помощью трофических пирамид чисел (численностей),
биомасс и энергий (рис. 3.3). Обычные пирамиды чисел, т.е. отображение
числа особей на каждом из трофических уровней данной экосистемы, для пастбищных (по Ю. Одуму.
1975) А - пирамида чисел, Б - пирамида биомасс, В - пирамида энергий. Данные
приведены в расчете на 4 га за год; шкалы логарифмические цепей имеют
широкое основание (большое число продуцентов, Р) и резкое сужение к конечным
консументам (рис. 3.3, А). При этом числа «ступеней» различаются не менее, чем
на 1 -3 порядка. Но это справедливо только для травяных сообществ - луговых или
степных биоценозов. Картина резко искажается, если рассматривать лесное
сообщество. На одном дереве могут кормиться тысячи фитофагов или на одном
трофическом уровне оказываются такие разные фитофаги, как тля или слон.

Рис. 3.3. Пример
простой трофической пирамиды
Это
искажение можно преодолеть с помощью пирамиды биомасс. В наземных
экосистемах биомасса растений всегда существенно больше биомассы животных, а
биомасса фитофагов всегда больше биомассы зоофагов (рис. 3.3, Б). Иначе
выглядят пирамиды биомасс для водных, особенно морских экосистем: биомасса
животных обычно намного больше биомассы растений. Эта «неправильность»
обусловлена тем, что пирамидами биомасс не учитывается продолжительность
существования поколений особей на разных трофических уровнях и скорость
образования и выеданйя биомассы. Главным продуцентом морских
экосистем является фитопланктон, имеющий большой репродуктивный потенциал и
быструю смену поколений. В океане за год может смениться до 50 поколений
фитопланктона. За то время, пока хищные рыбы (а тем более моржи и киты) накопят
свою биомассу, сменится множество поколений фитопланктона, суммарная биомасса
которых намного больше. Вот почему универсальным способом выражения трофической
структуры экосистем являются пирамиды скоростей образования живого вещества,
т.е. продуктивности. Их обычно называют пирамидами энергий, имея в вицу
энергетическое выражение продукции, хотя правильнее было бы говорить о
мощности.
Стабильность и развитие экосистем. В природных экосистемах происходят постоянные
изменения состояния популяций организмов. Они вызываются разными причинами.
Кратковременные - погодными условиями и биотическими воздействиями; сезонные
(особенно в умеренных и высоких широтах) - большим годовым ходом температуры;
от года к году - различными случайными сочетаниями абиотических и биотических
факторов. Однако все эти колебания, как правило, более или менее регулярны и не
выходят за границы устойчивости экосистемы - ее обычного размера, видового
состава, биомассы, продуктивности, соответствующих географическим и
климатическим условиям местности. Такое состояние экосистемы называется климаксным.
Климаксные
сообщества характеризуются устойчивым динамическим равновесием между
биотическими потенциалами входящих в сообщество популяций и сопротивлением
среды. Постоянство важнейших экологических параметров обозначают как гомеостаз
экосистемы. Устойчивость экосистемы тем. больше, чем больше она по размеру и
чем богаче и разнообразнее ее видовой и популяционный состав.
Стремясь
к поддержанию гомеостаза, экосистемы, тем не менее, способны к изменениям, к
развитию, к переходу от более простых к более сложным формам. Масштабные
изменения географической обстановки или типа ландшафта под влиянием природных
катастроф или деятельности человека приводят к определенным последовательным
изменениям состояния биогеоценозов местности - сукцессиям (от англ.
succession - последовательность).
Различают
первичную сукцессию - постепенное заселение организмами появившейся
девственной суши, оголенной материнской породы (отступившее море или ледник,
голые скалы и застывшая лава после вулканического извержения и т.п.). В этих
случаях решающую роль играет процесс почвообразования. Начальное выветривание -
разрушение и разрыхление поверхности минеральной основы под действием перепадов
температуры и увлажнения - уже может быть использовано бактериями, лишайниками,
а затем и редкой одноярусной пионерной растительностью. Ее появление, а с нею -
симбиотрофов и мелких животных значительно ускоряет образование почвы и постепенное
заселение территории сериями все более сложных растительных сообществ, все
более крупными растениями и животными. Так система постепенно проходит все
стадии развития до климаксного состояния.
Вторичные
сукцессии имеют характер постепенного
восстановления свойственного данной местности сообщества после нанесенных
повреждений (последствий бури, пожара, вырубки, наводнения, выпаса скота,
запуска полей). Возникшая в результате вторичной сукцессии климаксная система
может существенно отличаться от первоначальной, если изменились некоторые
характеристики ландшафта или климатические условия. Сукцессии происходят путем
замещения одних видов другими и поэтому их нельзя приравнивать к реакциям
гомеостаза.
Биомы. Наземные
экосистемы, относящиеся к одной природно-климатической зоне имеют общую
структуру доминирующей растительности и поэтому могут рассматриваться как
единый большой биогеоценоз - биом. Биомы являются основными объектами
экологической географии. Они различаются по климату, по многим особенностям
флоры и фауны, по биологическому разнообразию, но в пределах каждого биома можно
встретить множество сходных по приспособлению форм животных и растений, хотя
происхождение их различно. В табл. 3.1 приведены ориентировочные данные о
площади, биомассе и годовой продуктивности крупнейших биомов, охватывающих всю
Землю. По ним можно судить не только о больших экологических различиях
природных зон, но и о степени давления человека на природу планеты. Еще совсем
недавно при составлении подобных таблиц и карт экологи пренебрегали участками
суши, преобразованными человеком. Обращает на себя внимание, что 94% всей
биомассы Земли сосредоточено в лесах, более половины - во влажных тропических
лесах. Обрабатываемые земли, составившие 10% площади суши, дают такую же долю
первичной продукции, уступая по продуктивности степям, которые вытеснены пашней.
Таблица 3.1
Площади, биомасса и продуктивность основных биомов Земли
|
Основные биомы, категория земель
|
Площадь, млн км
|
Биомасса
(сухое вещество)
|
Готовая продукция
|
|
т/га
|
млрд т.
|
т/га
|
млрд т.
|
|
Тундры
и лесотундры
|
4,2
|
9
|
4
|
5
|
2,1
|
|
Таежные
и горные хвойные леса
|
12,8
|
227
|
290
|
9
|
11,5
|
|
Лиственно-хвойные
бореальные леса
|
6,2
|
280
|
174
|
14
|
8,6
|
|
Широколиственные
листопадные леса
|
7,6
|
325
|
248
|
15
|
11,4
|
|
Субтропические
леса
|
5,3
|
482
|
255
|
21
|
11,3
|
|
Влажные
тропические леса
|
10,3
|
960
|
990
|
36
|
37,1
|
|
Саванна,
чапаррель
|
6,2
|
100
|
62
|
15
|
9.2
|
|
Степи,
прерии
|
2,8
|
26
|
7
|
13
|
3,6
|
|
Пустыни
|
22,7
|
7
|
16
|
2
|
4,2
|
|
Пашня,
обрабатываемые земли
|
15,1
|
26
|
39
|
12
|
21,1 .
|
|
Освоенные
и окультуренные пастбища
|
26,3
|
16
|
42
|
7
|
18,3
|
|
Воды
суши
|
2,4
|
5
|
1
|
2
|
0,5
|
|
Сооружения,
дороги, горные выработки
|
9,8
|
|
|
|
|
|
Полярные
и горные льды
|
17,2
|
|
|
|
|
|
Итого
для всей суши
|
148,9
|
|
2128
|
|
139
|
|
Океан
|
361,1
|
|
7
|
|
80
|
|
Всего
|
510,0
|
|
2135
|
|
219
|
В
тех сообществах, где преобладают однолетние растения, травы (тундра, степи,
обрабатываемые земли), годовая продукция мало отличается от среднегодовой биомассы.
В лесах же продукция составляет лишь от 3,5 до 6,7% от биомассы. В океане
продукция заметно превышает среднегодовую биомассу.
|
